
Presumptive First Record of Myotis aurascens (Chiroptera, Vespertilionidae) from China with a Phylogenetic Analysis

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. MtDNA Sequencing, Assembly, and Annotation
2.3. Characteristic Analysis of Comparative Genomes
2.4. Phylogenetic Analysis
3. Results
3.1. Habitat and Morphology
3.2. Genome Organization
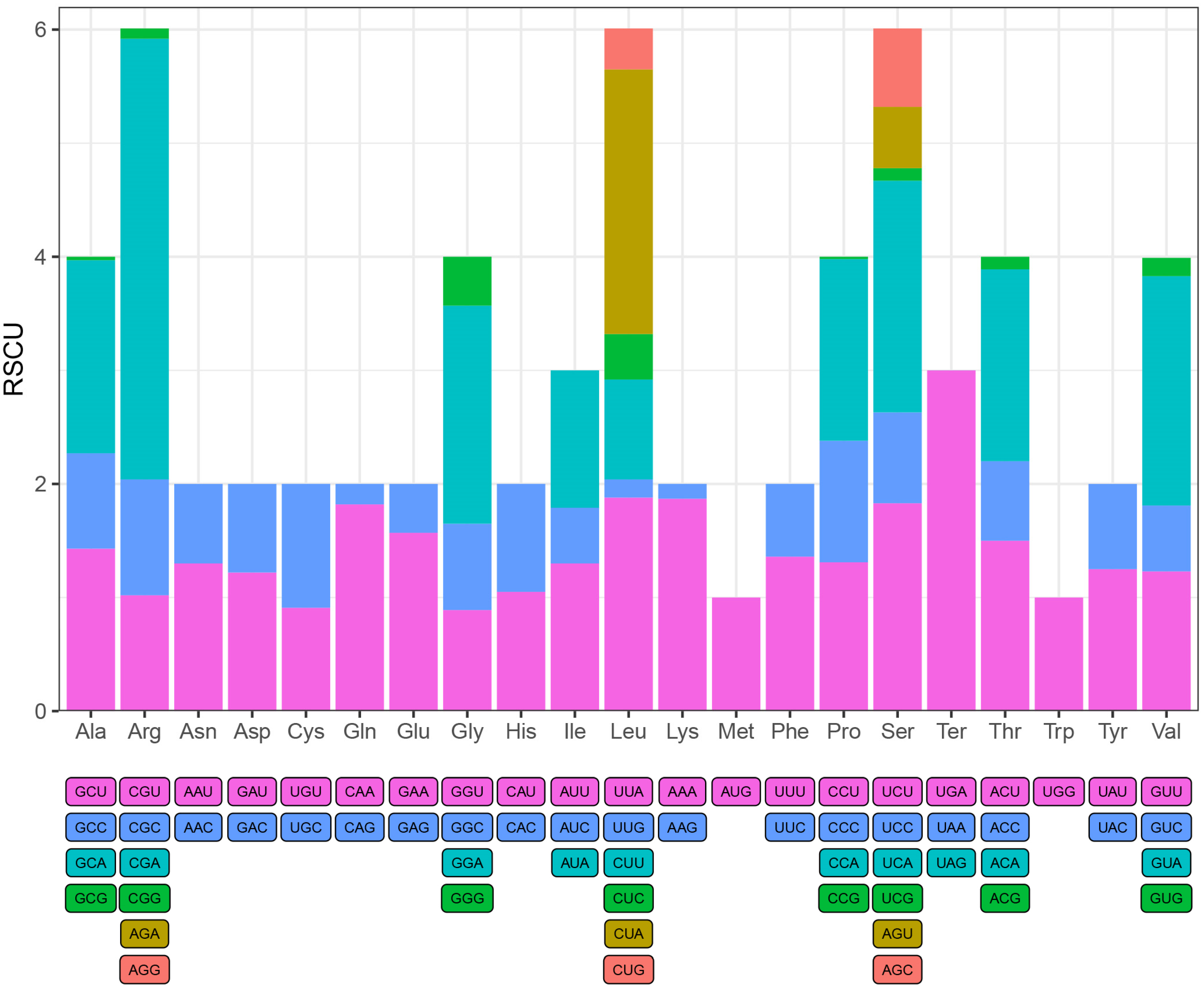
3.3. Characterization of Coding Genes
3.4. Phylogenetic Analysis within Myotis
3.5. Phylogenetic Analysis within Vespertilionidae
3.6. Genetic Distance Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bickham, J.W.; Patton, J.C.; Schlitter, D.A.; Rautenbach, I.L.; Honeycutt, R.L. Molecular phylogenetics, karyotypic diversity, and partition of the genus Myotis (Chiroptera: Vespertilionidae). Mol. Phylogenet. Evol. 2004, 33, 333–338. [Google Scholar] [CrossRef]
- Wilson, D.E.; Mittermeier, R.A. Handbook of the Mammals of the World (Volume 9); Lynx Edicions: Sardañola del Vallés, Spain, 2019. [Google Scholar]
- Wei, F.; Yang, Q.; Wu, Y.; Jiang, X.; Liu, S.; Li, B.; Yang, G.; Li, M.; Zhou, J.; Li, S. Catalogue of mammals in China (2021). Acta Theriol. Sin. 2021, 41, 487–501. [Google Scholar]
- Benda, P.; Paunović, M. Myotis aurascens. The IUCN Red List of Threatened Species. 2016. Available online: http://dx.doi.org/10.2305/IUCN.UK.2016-2.RLTS.T136553A21993953.en (accessed on 16 April 2022).
- Benda, P. First record of Myotis aurascens and second record of Myotis brandtii in Montenegro. LYNX 2004, 35, 13–18. [Google Scholar]
- Kim, Y.K.; Park, S.; Kim, T.W.; Park, J.H.; Adhikari, P.; Kim, G.R.; Park, S.M.; Lee, J.W.; Han, S.H.; Oh, H.S. Distribution Status and Phylogenetic Relationship of Myotis aurascens in Jeju Island. J. Life Sci. 2015, 25, 1354–1361. [Google Scholar] [CrossRef]
- Tsytsulina, K.; Dick, M.H.; Maeda, K.; Masuda, R. Systematics and phylogeography of the steppe whiskered bat Myotis aurascens Kuzyakin, 1935 (Chiroptera, Vespertilionidae). Russ. J. Theriol. 2012, 11, 1–20. [Google Scholar] [CrossRef]
- Benda, P. Myotis aurascens Kusjakin, 1935–Steppen-Bartfledermaus; Aula-Verlag: Wiebelsheim, Germany, 2004; pp. 1149–1158. [Google Scholar]
- Li, H.; Kong, L.; Wang, K.; Zhang, S.; Motokawa, M.; Wu, Y.; Wang, W.; Li, Y. Molecular phylogeographic analyses and species delimitations reveal that Leopoldamys edwardsi (Rodentia: Muridae) is a species complex. Integr. Zool. 2019, 14, 494–505. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Li, H.; Wang, J.; Li, Y. Niviventer confucianus sacer (Rodentia, Muridae) is a distinct species based on molecular, karyotyping, and morphological evidence. ZooKeys 2020, 959, 137–159. [Google Scholar] [CrossRef]
- Lu, L.; Ge, D.; Chesters, D.; Ho, S.Y.W.; Ma, Y.; Li, G.; Wen, Z.; Wu, Y.; Wang, J.; Xia, L.; et al. Molecular phylogeny and the underestimated species diversity of the endemic white-bellied rat (Rodentia: Muridae: Niviventer) in Southeast Asia and China. Zool. Scr. 2015, 44, 475–494. [Google Scholar] [CrossRef]
- Mori, S.; Matsunami, M. Signature of positive selection in mitochondrial DNA in Cetartiodactyla. Genes Genet. Syst. 2018, 93, 65–73. [Google Scholar] [CrossRef]
- Pan, T.; Sun, Z.; Lai, X.; Orozcoterwengel, P.; Yan, P.; Wu, G.; Wang, H.; Zhu, W.; Wu, X.; Zhang, B. Hidden species diversity in Pachyhynobius: A multiple approaches species delimitation with mitogenomes. Mol. Phylogenet. Evol. 2019, 137, 138–145. [Google Scholar] [CrossRef]
- Yang, X.; Wang, Q.; Zhang, L.; Bao, S.; Zhao, S.; Dou, H.; Zhang, H. The complete mitochondrial genome of Steppe Whiskered Bat (Myotis aurascens; Kuzyakin, 1935) and phylogenetic analysis. Mitochondrial DNA Part B 2022, 7, 611–612. [Google Scholar] [CrossRef] [PubMed]
- Coil, D.; Jospin, G.; Darling, A.E. A5-miseq: An updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 2015, 31, 587–589. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Earl, A.M. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Alzohairy, A.M. BioEdit: An important software for molecular biology. Gerf Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Posada, D. Model Test: A Tool to Select the Best-Fit Model of Nucleotide Substitution. 2005. Available online: https://evomics.org/resources/software/molecular-evolution-software/modeltest/ (accessed on 1 April 2022).
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2013, 9, 722. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Swofford, D. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods). 2002. Available online: http://phylosolutions.com/paup-test/ (accessed on 1 April 2022).
- Jolle, B.S.; Veronika, B.; Du, P.L.; Denise, K.; Carsten, M.; Venelin, M.; Müller, N.F.; Jūlija, P.; Rasmussen, D.A.; Chi, Z. Taming the BEAST—A Community Teaching Material Resource for BEAST 2. Syst. Biol. 2018, 67, 170–174. [Google Scholar]
- Sudhir, K.; Glen, S.; Koichiro, T. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar]
- Oh, D. Geographical Distribution of Korean Bats and Morphology of Its Skull. Master’s Thesis, Incheon National University, Incheon, Republic of Korea, 2013. [Google Scholar]
- Ren, R.; Shi, S.; Wu, Q.; Deng, X.; Chen, Y. David’s myotis (Myotis davidii) found in Hengdong County, Hunan Province, China. Chin. J. Zool. 2017, 53, 870–876. [Google Scholar]
- Jebb, D.; Foley, N.M.; Kerth, G.; Teeling, E.C. The complete mitochondrial genome of the Bechstein’s bat, Myotis bechsteinii (Chiroptera, Vespertilionidae). Mitochondrial DNA Part B 2017, 2, 92–94. [Google Scholar] [CrossRef]
- Wei, S.J.; Shi, M.; Chen, X.X.; Sharkey, M.J.; van Achterberg, C.; Ye, G.Y.; He, J.H. New views on strand asymmetry in insect mitochondrial genomes. PLoS ONE 2010, 5, e12708. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, M.; Chen, Y.; Shen, Q.Q.; Sun, Y.X.; Zhang, L.B. Myotis nipalensis Discovered in Hubei and Jiangsu Provinces, China. Chin. J. Zool. 2014, 49, 483–489. [Google Scholar]
- Smith, A.T.; Xie, Y.; Hoffmann, R.S.; Lunde, D.; MacKinnon, J.; Wilson, D.E.; Wozencraft, W.C. A Guide to the Mammals of China; Princeton University Press: Princeton, NJ, USA, 2010. [Google Scholar]
- Wang, Y. A Complete Checklist of Mammal Species and Subspecies in China a Taxonomic and Geographic Reference; China Forestry Publishing House: Beijing, China, 2003. [Google Scholar]
- Benda, P.; Tsytsulina, K. Taxonomic revision of Myotis mystacinus group (Mammalia: Chiroptera). Acta Soc. Zool. Bohem. 2000, 64, 331–398. [Google Scholar]
- Chung, C.U.; Kim, S.C.; Sang, H.H. Diurnal Roosts Selection and Home Range Size in the Myotis aurascens (Chiroptera: Vespertilionidae) Inhabiting a Rural Area. J. Environ. Sci. Int. 2013, 22, 1227–1234. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement Index (g, mm) | M. aurascens | M. davidii | ||
|---|---|---|---|---|
| n = 1 (This Study) | n = 10 [6] | n = 7 [27] | n = 3 * [28] | |
| Body mass (BM) | 6.33 | 5.70–7.10 | - | 4.50–5.00 |
| Forearm length (FL) | 35.87 | 34.71–38.21 | 33.80–37.30 | 34.00–34.54 |
| Length of head and body (LHB) | 45.10 | 42.64–46.40 | 40.80–48.00 | 38.00–41.00 |
| Tibia length (TBL) | 15.11 | 16.22–18.04 | 15.10–17.47 | 15.00–15.11 |
| Tail length (TL) | 32.70 | 41.03–45.26 | 36.00–42.00 | 31.00–35.00 |
| Ear length (EL) | 13.22 | 13.06–14.98 | 11.00–15.00 | 11.00–15.00 |
| Ear width (EW) | 7.25 | - | - | 5.51 |
| Tragus length (TRL) | 7.51 | 7.64–8.88 | 6.30–9.00 | 5.09 |
| Length of hindfoot (LHF) | 7.15 | 6.42–7.68 | 7.00–10.00 | 7.62–9.50 |
| Wing length (WL) | 91.67 | - | - | - |
| Wing span (WS) | 216.44 | - | - | - |
| Third metacarpal length (Mc III) | 28.31 | - | - | 31.94 |
| Length of first phalanx of the third digit (III1) | 10.27 | - | - | 12.02 |
| Length of second phalanx of the third digit III2) | 15.55 | - | - | 9.06 |
| Fourth metacarpal length (Mc IV) | 27.80 | - | - | 30.08 |
| Length of first phalanx of the fourth digit IV1) | 7.85 | - | - | - |
| Length of second phalanx of the fourth digit (IV2) | 7.49 | - | - | - |
| Fifth metacarpal length (Mc V) | 29.55 | - | - | 32.67 |
| Length of first phalanx of the fifth digit (V1) | 16.46 | - | - | 8.37 |
| Gene | Strand | Location | Size (bp) | Base Composition (%) | Start Codon | Stop Codon | |||
|---|---|---|---|---|---|---|---|---|---|
| A(U) | C | G | T | ||||||
| tRNAPhe | H | 1–68 | 68 | 36.76 | 22.06 | 16.18 | 25.00 | - | - |
| 12S rRNA | H | 69–1034 | 966 | 36.96 | 21.22 | 17.49 | 24.33 | - | - |
| tRNAVal | H | 1035–1103 | 69 | 39.13 | 17.39 | 13.04 | 30.43 | - | - |
| 16S rRNA | H | 1103–2670 | 1568 | 38.90 | 18.56 | 15.68 | 25.96 | - | - |
| tRNALeu2 | H | 2672–2746 | 75 | 30.67 | 18.67 | 21.33 | 29.33 | - | - |
| ND1 | H | 2752–3707 | 956 | 32.74 | 23.85 | 11.09 | 32.32 | ATG | TA- |
| tRNAIle | H | 3708–3776 | 69 | 34.78 | 11.59 | 17.39 | 36.23 | - | - |
| tRNAGln | L | 3774–3847 | 74 | 29.73 | 10.81 | 24.32 | 35.14 | - | - |
| tRNAMet | H | 3848–3916 | 69 | 28.99 | 24.64 | 18.84 | 27.54 | - | - |
| ND2 | H | 3917–4958 | 1042 | 39.25 | 25.24 | 7.58 | 27.93 | ATT | T-- |
| tRNATrp | H | 4959–5026 | 68 | 33.82 | 22.06 | 16.18 | 27.94 | - | - |
| tRNAAla | L | 5032–5100 | 69 | 28.99 | 11.59 | 21.74 | 37.68 | - | - |
| tRNAAsn | L | 5102–5174 | 73 | 24.66 | 13.70 | 21.92 | 39.73 | - | - |
| tRNACys | L | 5207–5272 | 66 | 27.27 | 21.21 | 24.24 | 27.27 | - | - |
| tRNATyr | L | 5273–5341 | 69 | 36.23 | 17.39 | 20.29 | 26.09 | - | - |
| COX1 | H | 5343–6887 | 1545 | 27.06 | 20.65 | 16.89 | 35.40 | ATG | TAA |
| tRNASer2 | L | 6901–6969 | 69 | 24.64 | 15.94 | 24.64 | 34.78 | - | - |
| tRNAAsp | H | 6977–7043 | 67 | 37.31 | 11.94 | 13.43 | 37.31 | - | - |
| COX2 | H | 7044–7727 | 684 | 33.92 | 21.93 | 12.57 | 31.58 | ATG | TAA |
| tRNALys | H | 7731–7798 | 68 | 35.29 | 19.12 | 14.71 | 30.88 | - | - |
| ATP8 | H | 7799–8002 | 204 | 40.22 | 22.55 | 6.86 | 30.39 | ATG | TAA |
| ATP6 | H | 7960–8640 | 681 | 32.60 | 23.05 | 11.60 | 32.75 | ATG | TAA |
| COX3 | H | 8640–9423 | 784 | 28.57 | 22.70 | 14.16 | 34.57 | ATG | T-- |
| tRNAGly | H | 9424–9493 | 70 | 37.14 | 17.14 | 14.29 | 31.43 | - | - |
| ND3 | H | 9494–9840 | 347 | 30.84 | 20.17 | 11.24 | 37.75 | ATA | TA- |
| tRNAArg | H | 9841–9909 | 69 | 42.03 | 8.70 | 8.70 | 40.58 | - | - |
| ND4L | H | 9911–10,207 | 297 | 28.28 | 22.56 | 12.12 | 37.04 | ATG | TAA |
| ND4 | H | 10,201–11,579 | 1379 | 33.14 | 23.42 | 10.51 | 32.92 | ATG | TA- |
| tRNAHis | H | 11,580–11,647 | 68 | 45.59 | 8.82 | 10.29 | 35.29 | - | - |
| tRNASer1 | H | 11,647–11,705 | 59 | 33.90 | 20.34 | 16.95 | 28.81 | - | - |
| tRNALeu1 | H | 11,706–11,775 | 70 | 38.57 | 14.29 | 20.00 | 27.14 | - | - |
| ND5 | H | 11,777–13,597 | 1821 | 33.44 | 22.73 | 10.16 | 33.66 | ATA | TAA |
| ND6 | L | 13,581–14,105 | 525 | 24.19 | 6.67 | 25.71 | 43.43 | ATA | TAA |
| tRNAGlu | L | 14,108–14,176 | 69 | 28.99 | 14.49 | 21.74 | 34.78 | - | - |
| Cytb | H | 14,184–15,323 | 1140 | 29.82 | 23.95 | 13.07 | 33.16 | ATG | AGA |
| tRNAThr | H | 15,324–15,393 | 70 | 37.14 | 14.29 | 18.57 | 30.00 | - | - |
| tRNAPro | L | 15,393–15,458 | 66 | 24.24 | 10.61 | 27.27 | 37.88 | - | - |
| D-loop | H | 15,459–16,771 | 1313 | 34.96 | 23.76 | 14.09 | 27.19 | - | - |
| Total | 16,771 | 33.93 | 22.17 | 12.96 | 30.94 | ||||
| Species | M. aurascens | M. brandtii | M. davidii | M. muricola | M. bombinus | M. frater | M. horsfieldii | M. myotis | M. pilosus |
|---|---|---|---|---|---|---|---|---|---|
| M. aurascens | - | 0.182 | 0.161 | 0.177 | 0.168 | 0.163 | 0.187 | 0.158 | 0.154 |
| M. brandtii | 0.152 | - | 0.195 | 0.213 | 0.200 | 0.199 | 0.214 | 0.193 | 0.19 |
| M. davidii | 0.138 | 0.161 | - | 0.188 | 0.178 | 0.168 | 0.176 | 0.163 | 0.163 |
| M. muricola | 0.149 | 0.172 | 0.156 | - | 0.192 | 0.186 | 0.205 | 0.186 | 0.181 |
| M. bombinus | 0.143 | 0.163 | 0.150 | 0.159 | - | 0.172 | 0.201 | 0.135 | 0.168 |
| M. frater | 0.140 | 0.164 | 0.143 | 0.155 | 0.146 | - | 0.198 | 0.163 | 0.164 |
| M. horsfieldii | 0.156 | 0.172 | 0.148 | 0.167 | 0.165 | 0.163 | - | 0.195 | 0.188 |
| M. myotis | 0.136 | 0.159 | 0.139 | 0.155 | 0.119 | 0.140 | 0.161 | - | 0.162 |
| M. pilosus | 0.133 | 0.157 | 0.140 | 0.151 | 0.143 | 0.140 | 0.156 | 0.139 | - |
| Species | M. aurascens | M. ikonnikovi | M. alcathoe | M. mystacinus | M. davidii | M. dasycneme | M. montivagus | M. muricola | M. daubentonii |
|---|---|---|---|---|---|---|---|---|---|
| M. aurascens | - | 0.126 | 0.148 | 0.129 | 0.156 | 0.154 | 0.174 | 0.169 | 0.168 |
| M. ikonnikovi | 0.112 | - | 0.16 | 0.138 | 0.165 | 0.151 | 0.17 | 0.167 | 0.172 |
| M. alcathoe | 0.128 | 0.137 | - | 0.088 | 0.166 | 0.174 | 0.179 | 0.183 | 0.172 |
| M. mystacinus | 0.114 | 0.120 | 0.080 | - | 0.176 | 0.176 | 0.189 | 0.18 | 0.172 |
| M. davidii | 0.134 | 0.140 | 0.140 | 0.147 | - | 0.168 | 0.17 | 0.184 | 0.171 |
| M. dasycneme | 0.132 | 0.129 | 0.147 | 0.147 | 0.142 | - | 0.174 | 0.174 | 0.175 |
| M. montivagus | 0.146 | 0.143 | 0.150 | 0.157 | 0.143 | 0.145 | - | 0.184 | 0.18 |
| M. muricola | 0.143 | 0.140 | 0.151 | 0.150 | 0.153 | 0.146 | 0.153 | - | 0.188 |
| M. daubentonii | 0.142 | 0.144 | 0.144 | 0.144 | 0.144 | 0.147 | 0.149 | 0.155 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Chen, X.; Gao, X.; Sun, G.; Song, X.; Dou, H.; Zhang, H. Presumptive First Record of Myotis aurascens (Chiroptera, Vespertilionidae) from China with a Phylogenetic Analysis. Animals 2023, 13, 1629. https://doi.org/10.3390/ani13101629
Yang X, Chen X, Gao X, Sun G, Song X, Dou H, Zhang H. Presumptive First Record of Myotis aurascens (Chiroptera, Vespertilionidae) from China with a Phylogenetic Analysis. Animals. 2023; 13(10):1629. https://doi.org/10.3390/ani13101629
Chicago/Turabian StyleYang, Xiufeng, Xingyao Chen, Xiaodong Gao, Guolei Sun, Xue Song, Huashan Dou, and Honghai Zhang. 2023. "Presumptive First Record of Myotis aurascens (Chiroptera, Vespertilionidae) from China with a Phylogenetic Analysis" Animals 13, no. 10: 1629. https://doi.org/10.3390/ani13101629