Listeria monocytogenes Infection of Bat Pipistrellus nathusii Epithelial cells Depends on the Invasion Factors InlA and InlB

 , ,
, ,  ,
,
Abstract
:
1. Introduction
2. Results
2.1. Primary Cell Cultures and Their Characteristics
2.2. L. monocytogenes Invades and Reproduces in Bat Kidney Cells
2.3. L. monocytogenes Cell-to-Cell Spread
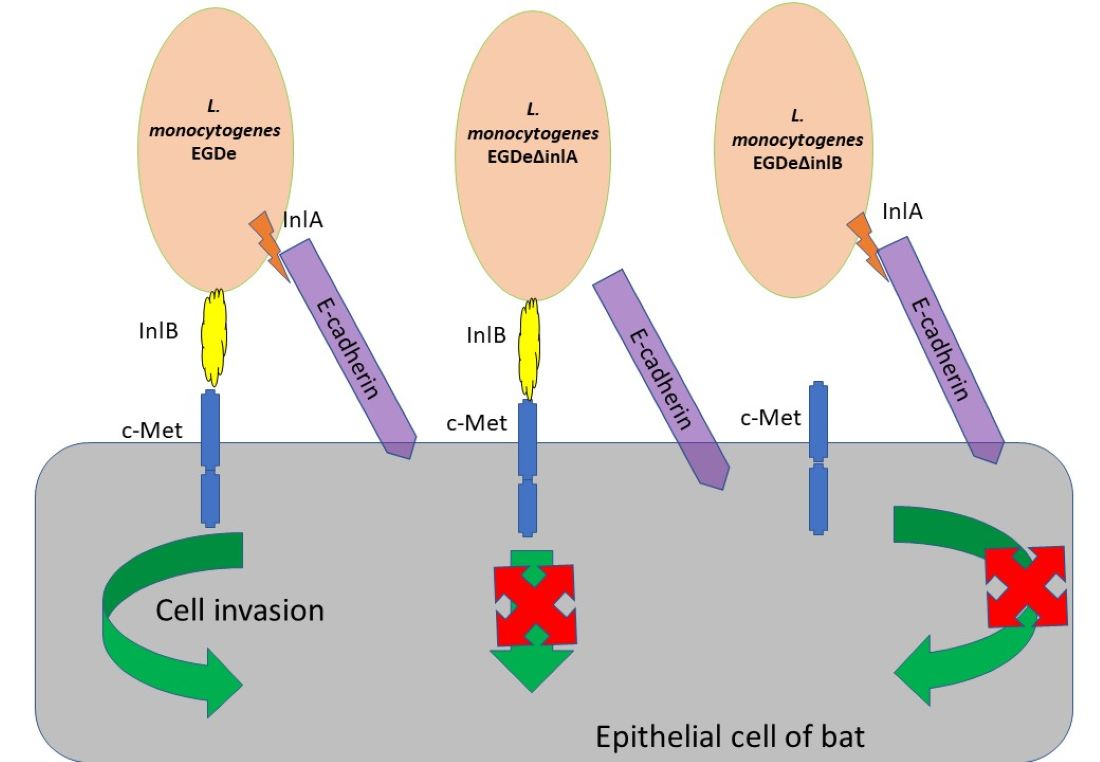
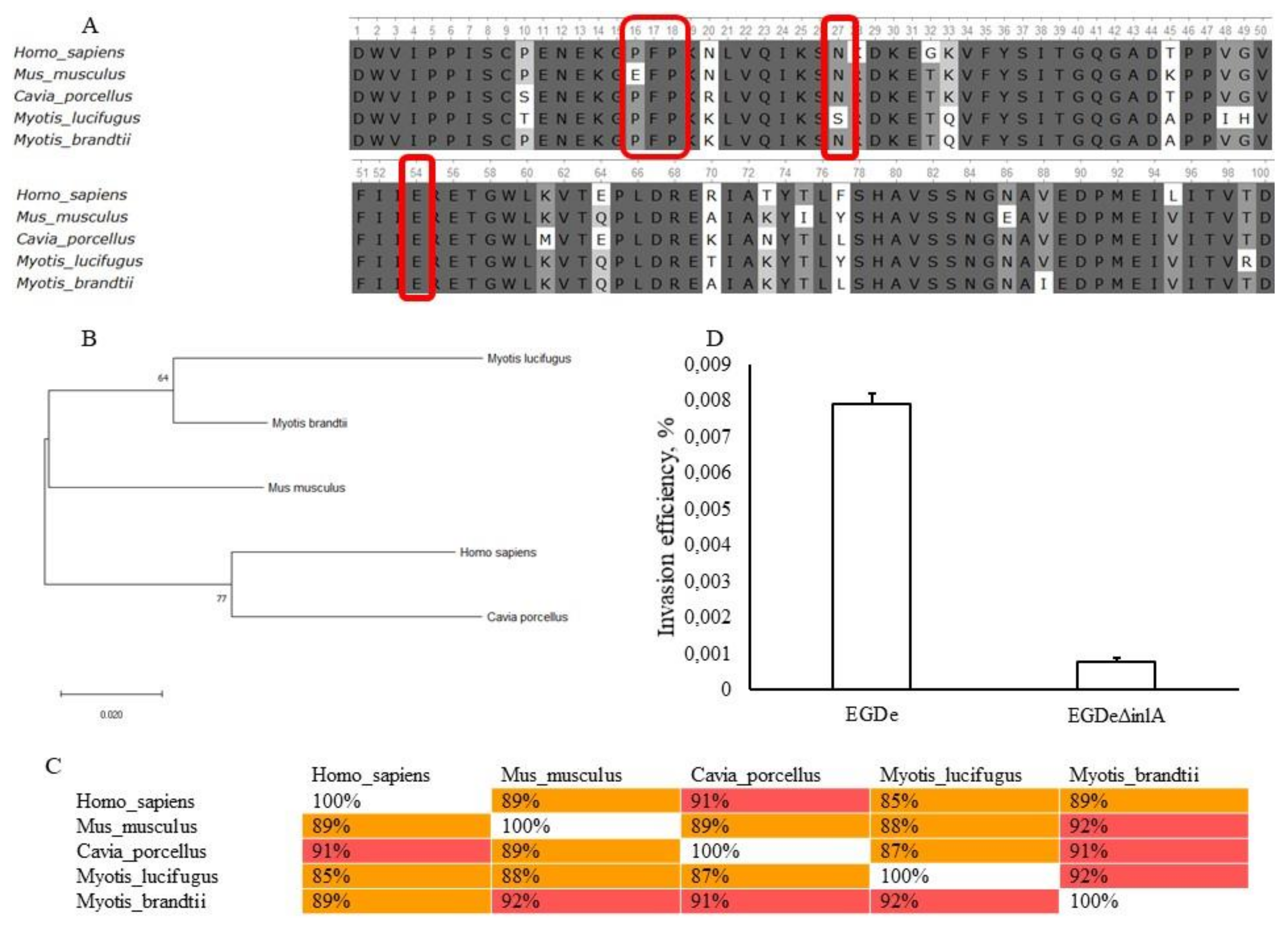
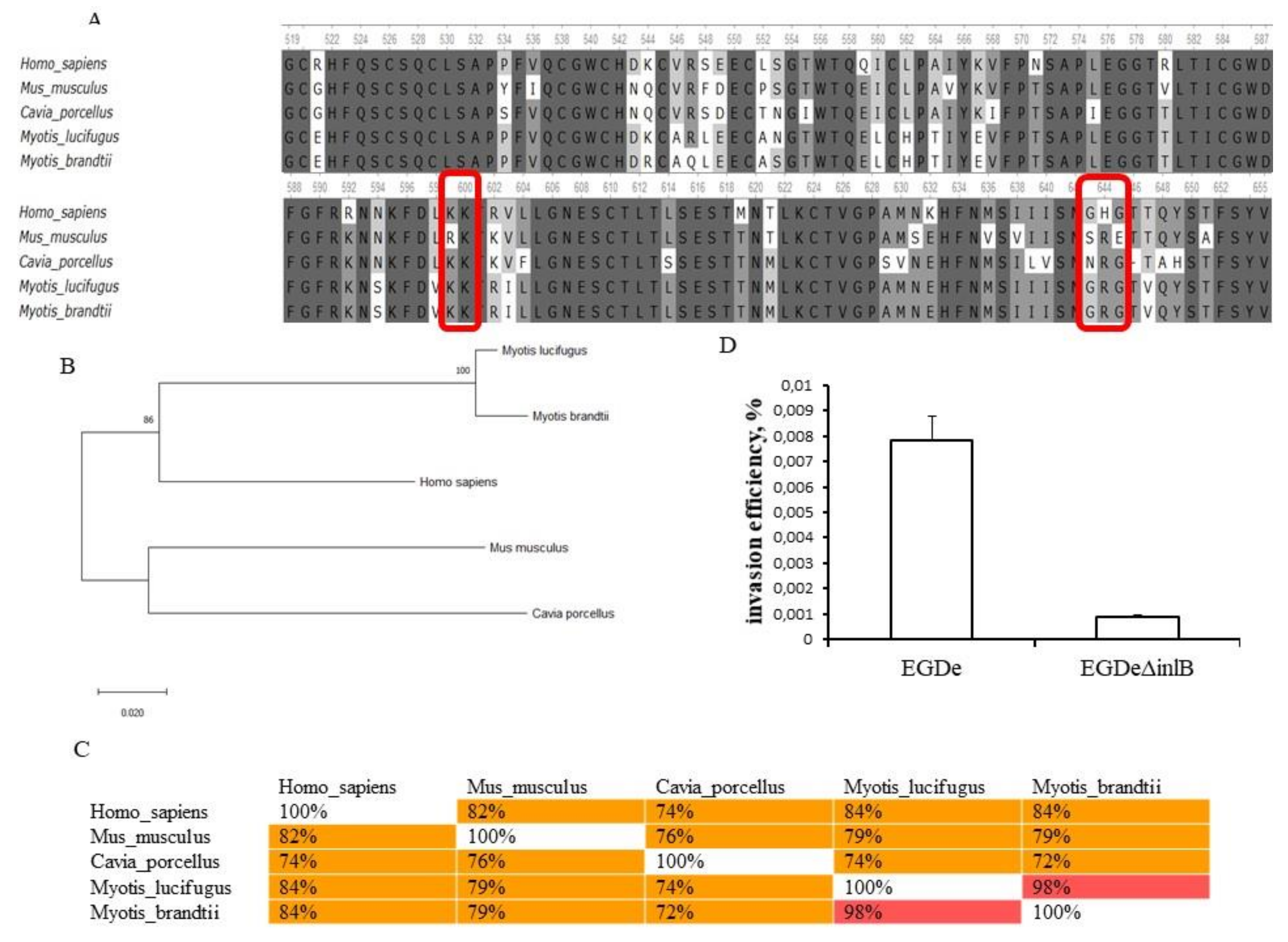
2.4. Invasion Factors InlA and InlB Are Required for L. monocytogenes Invasion into bKEC
2.5. Natural InlB Variants Differently Restored Invasion of the Strain EGDeΔinlB.
3. Discussion
4. Materials and methods
4.1. Animals
4.2. Tissue Trypsinization and Cell Isolation
4.3. Cell Culture and Subculture
4.4. Cryopreservation and Thawing of Cells
4.5. Chromosome Preparations and Karyological Analysis
4.6. Bacterial Strains and Growth Conditions
4.7. In Vitro Invasion and Proliferation Assay
4.8. Sequence Analysis
4.9. Immunoblotting
4.10. Primary Antibody Development and Purification
4.11. Plaque Forming Assay
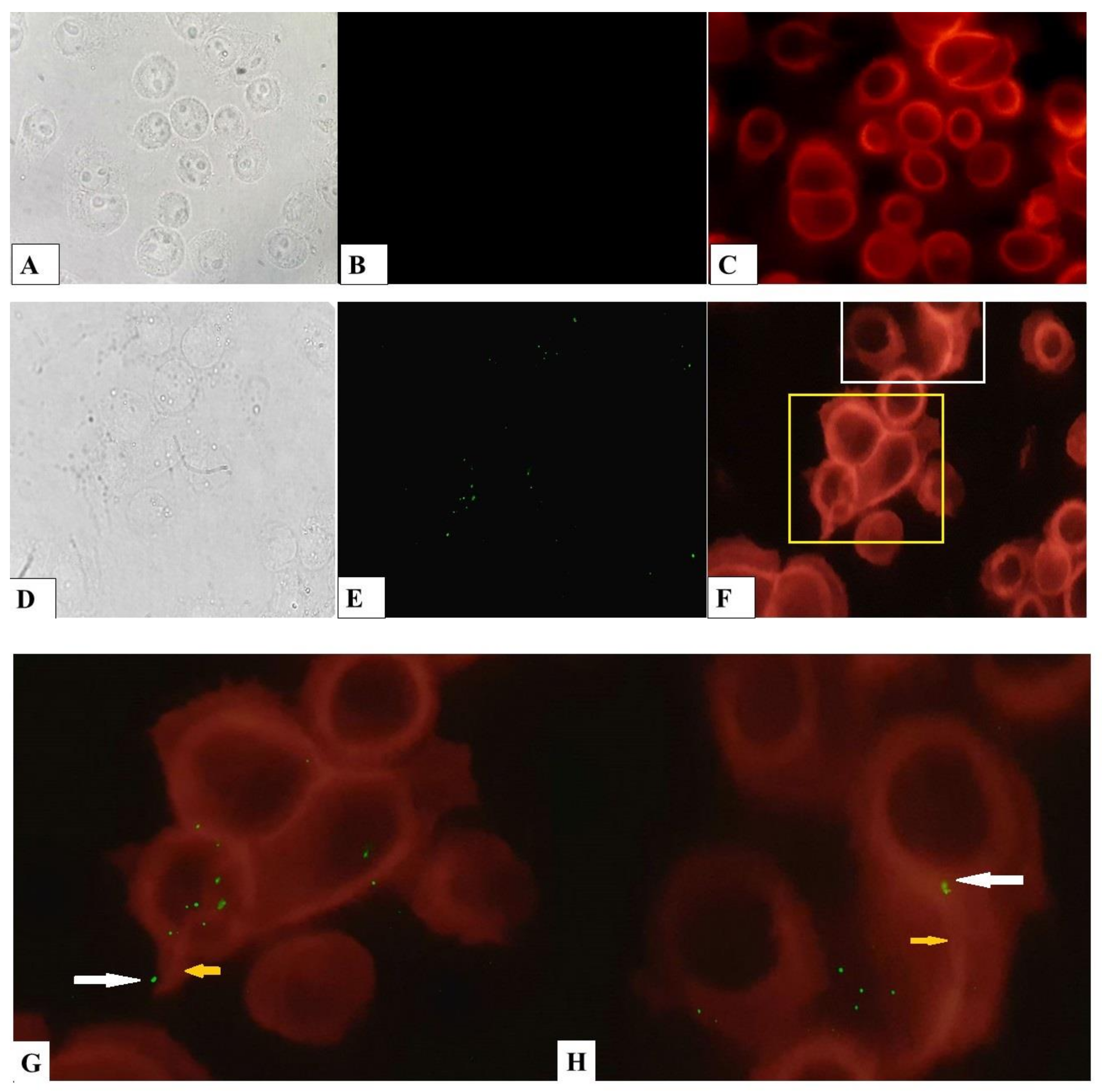
4.12. Determination of Cytoskeleton Rearrangements and Bacterial Cells Visualization in bKEC
4.13. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brook, C.E.; Dobson, A.P. Bats as “special” reservoirs for emerging zoonotic pathogens. Trends Microbiol. 2015, 23, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Kohl, C.; Kurth, A. European Bats as Carriers of Viruses with Zoonotic Potential. Viruses 2014, 6, 3110–3128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, G.; Wang, Q.; Gao, G.F. Bat-to-human: Spike features determining “host jump” of coronaviruses SARS-CoV, MERS-CoV, and beyond. Trends Microbiol. 2015, 23, 468–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneeberger, K.; Voigt, C.C. Zoonotic Viruses and Conservation of Bats. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Springer International Publishing: Cham, Switzerland, 2016; pp. 263–292. [Google Scholar]
- Wang, L.-F.; Anderson, D.E. Viruses in bats and potential spillover to animals and humans. Curr. Opin. Virol. 2019, 34, 79–94. [Google Scholar] [CrossRef]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.; Lau, S.; Woo, P.; Yuen, K.Y. Bats as a continuing source of emerging infections in humans. Rev. Med. Virol. 2007, 17, 67–91. [Google Scholar] [CrossRef]
- Mühldorfer, K. Bats and bacterial pathogens: A review. Zoonoses Public Health 2013, 60, 93–103. [Google Scholar] [CrossRef]
- Banskar, S.; Bhute, S.S.; Suryavanshi, M.; Punekar, S.; Shouche, Y.S. Microbiome analysis reveals the abundance of bacterial pathogens in Rousettus leschenaultii guano. Sci. Rep. 2016, 6, 36948. [Google Scholar] [CrossRef]
- Adesiyun, A.A.; Stewart-Johnson, A.; Thompson, N.N. Isolation of enteric pathogens from bats in trinidad. J. Wildl. Dis. 2009, 45, 952–961. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.M.; Kloepper, L.N.; Duncan, M.; McInroy, J.A.; Kloepper, J.W. Variation in Bat Guano Bacterial Community Composition With Depth. Front. Microbiol. 2018, 9, 914. [Google Scholar] [CrossRef]
- Childs-Sanford, S.E.; Kollias, G.V.; Abou-Madi, N.; McDonough, P.L.; Garner, M.M.; Mohammed, H.O. Yersinia pseudotuberculosis in a closed colony of Egyptian fruit bats (Rousettus aegyptiacus). J. Zoo Wildl. Med. 2009, 40, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.; Zarnegar, R.; Michalopoulos, G.K. Characterization of the effects of human placental HGF on rat hepatocytes. J. Cell. Physiol. 1992, 150, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Höhne, K.; Loose, B.; Seeliger, H.P. Isolation of Listeria monocytogenes in slaughter animals and bats of Togo (West Africa). Ann. Microbiol. (Paris) 1975, 126A, 501–507. [Google Scholar]
- Allocati, N.; Petrucci, A.G.; Di Giovanni, P.; Masulli, M.; Di Ilio, C.; De Laurenzi, V. Bat–man disease transmission: Zoonotic pathogens from wildlife reservoirs to human populations. Cell Death Discov. 2016, 2, 16048. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Settai, S.; Hayashidani, H.; Urabe, T.; Namai, S.; Une, Y. Outbreak of Yersiniosis in Egyptian Rousette Bats (Rousettus aegyptiacus) Caused by Yersinia pseudotuberculosis Serotype 4b. J. Comp. Pathol. 2013, 148, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.H. A review of pseudotuberculosis at a European zoo: Epidemiology and approaches to control. In Proceedings of the American Association of Zoo Veterinarians; Health and Conservation of Captive and Free-Ranging Wildlife, San-Diego, CA, USA, 28 August–2 September 2004; 2004; pp. 303–309. [Google Scholar]
- Schipper, J.; Chanson, J.S.; Chiozza, F.; Cox, N.A.; Hoffmann, M.; Katariya, V.; Lamoreux, J.; Rodrigues, A.S.L.; Stuart, S.N.; Temple, H.J.; et al. The status of the world’s land and marine mammals: Diversity, threat, and knowledge. Science (80-) 2008, 322, 225–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clawson, R.L. Trends in population size and current status. In The Indiana Bat: Biology and Management of an Endangeredspecies; Kurta, A., Kennedy, J., Eds.; BatConservation International: Austin, TX, USA, 2002; pp. 2–8. [Google Scholar]
- Horn, J.W.; Arnett, E.B.; Kunz, T.H. Behavioral Responses of Bats to Operating Wind Turbines. J. Wildl. Manag. 2008, 72, 123–132. [Google Scholar] [CrossRef]
- Kunz T., H.; Reynolds, D.S. Bat colonies in buildings. In Monitoring Bat Populations in the United States and Territories: Problems and Prospects; O’Shea, T.J., Bogan, M.A., Eds.; US Geological Survey Information and Technology Report ITR 2003–003: Washington, DC, USA, 2003; pp. 91–102. [Google Scholar]
- McCracken, G.F. Estimates of population sizes in summer colonies of Brazilian free-tailed bats (Tadarida brasiliensis). In Monitoring Bat Populations in the United States and Territories: Problems and Prospects; O’Shea, T.J., Bogan, M.A., Eds.; US Geological Survey Information and Technology Report ITR 2003–003: Washington, DC, USA, 2003; pp. 21–30. [Google Scholar]
- Sgro, M.P.; Wilkins, K.T. Roosting behavior of the Mexican free-tailed bat (Tadarida brasiliensis) in a highway overpass. West. N. Am. Nat. 2003, 63, 366–373. [Google Scholar]
- Vázquez-Boland, J.A.; Kuhn, M.; Berche, P.; Chakraborty, T.; Domínguez-Bernal, G.; Goebel, W.; González-Zorn, B.; Wehland, J.; Kreft, J. Listeria pathogenesis and molecular virulence determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef] [Green Version]
- Weis, J.; Seeliger, H.P. Incidence of Listeria monocytogenes in nature. Appl. Microbiol. 1975, 30, 29–32. [Google Scholar] [CrossRef] [Green Version]
- Zaytseva, E.; Ermolaeva, S.; Somov, G.P. Low genetic diversity and epidemiological significance of Listeria monocytogenes isolated from wild animals in the far east of Russia. Infect. Genet. Evol. 2007, 7, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Voronina, O.L.; Ryzhova, N.N.; Kunda, M.S.; Kurnaeva, M.A.; Semenov, A.N.; Aksenova, E.I.; Egorova, I.Y.; Kolbasov, D.V.; Ermolaeva, S.A.; Gintsburg, A.L. Diversity and Pathogenic Potential of Listeria monocytogenes Isolated from Environmental Sources in the Russian Federation. Int. J. Mod. Eng. Res. 2015, 5, 5–15. [Google Scholar]
- Nilsson, A.; Karlsson, K. Listeria monocytogenes isolations from animals in Sweden during 1948 to 1957. Nord. Vet. Med 1959, 11, 305–315. [Google Scholar]
- Hayashidani, H.; Kanzaki, N.; Kaneko, Y.; Okatani, A.T.; Taniguchi, T.; Kaneko, K.I.; Ogawa, M. Occurrence of Yersiniosis and listeriosis in wild boars in Japan. J. Wildl. Dis. 2002, 38, 202–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adgamov, R.; Zaytseva, E.; Thiberge, J.-M.; Brisse, S.; Ermolaev, S. Genetically Related Listeria Monocytogenes Strains Isolated from Lethal Human Cases and Wild Animals. In Genetic Diversity in Microorganisms; IntechOpen: London, UK, 2012; pp. 235–250. [Google Scholar]
- Wiedmann, M. ADSA Foundation Scholar Award—An Integrated Science-Based Approach to Dairy Food Safety: Listeria monocytogenes as a Model System. J. Dairy Sci. 2003, 86, 1865–1875. [Google Scholar] [CrossRef]
- Eliott, D.; O’Brien, T.P.; Green, W.R.; Jampel, H.D.; Goldberg, M.F. Elevated intraocular pressure, pigment dispersion and dark hypopyon in endogenous endophthalmitis from Listeria monocytogenes. Surv. Ophthalmol. 1992, 37, 117–124. [Google Scholar] [CrossRef]
- Zaidman, G.W.; Coudron, P.; Piros, J. Listeria monocytogenes keratitis. Am. J. Ophthalmol. 1990, 109, 334–339. [Google Scholar] [CrossRef]
- Erdogan, H.M. Listerial Keratoconjunctivitis and Uveitis (Silage Eye). Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 505–510. [Google Scholar] [CrossRef]
- Morgand, M.; Leclercq, A.; Maury, M.M.; Bracq-Dieye, H.; Thouvenot, P.; Vales, G.; Lecuit, M.; Charlier, C. Listeria monocytogenes-associated respiratory infections: A study of 38 consecutive cases. Clin. Microbiol. Infect. 2018, 24, 1339.e1–1339.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelenik, K.; Avberšek, J.; Pate, M.; Lušicky, M.; Krt, B.; Ocepek, M.; Zdovc, I. Cutaneous Listeriosis in a Veterinarian with the Evidence of Zoonotic Transmission—A Case Report. Zoonoses Public Health 2014, 61, 238–241. [Google Scholar] [CrossRef]
- Regan, E.J.; Harrison, G.A.J.; Butler, S.; McLauchlin, J.; Thomas, M.; Mitchell, S. Primary cutaneous listeriosis in a veterinarian. Vet. Rec. 2005, 157, 207. [Google Scholar] [CrossRef]
- Godshall, C.E.; Suh, G.; Lorber, B. Cutaneous listeriosis. J. Clin. Microbiol. 2013, 51, 3591–3596. [Google Scholar] [CrossRef] [Green Version]
- Bakulov, I.A.; Vasylyev, D.A.; Kolbasov, D.B.; Kovaleva, E.N.; Egorova, I.Y.; Selyaniniov, Y.O. Listeria and Listeriosis, 2nd ed.; RDICMB: Ulyanovsk, Russia, 2016. [Google Scholar]
- Lecuit, M. Understanding how Listeria monocytogenes targets and crosses host barriers. Clin. Microbiol. Infect. 2005, 11, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Lingnau, A.; Domann, E.; Hudel, M.; Bock, M.; Nichterlein, T.; Wehland, J.; Chakraborty, T. Expression of the Listeria monocytogenes EGD inlA and inlB genes, whose products mediate bacterial entry into tissue culture cell lines, by PrfA-dependent and -independent mechanisms. Infect. Immun. 1995, 63, 3896–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bierne, H.; Cossart, P. InlB, a surface protein of Listeria monocytogenes that behaves as an invasin and a growth factor. J. Cell Sci. 2002, 115, 3357–3367. [Google Scholar] [PubMed]
- Lecuit, M.; Ohayon, H.; Braun, L.; Mengaud, J.; Cossart, P. Internalin of Listeria monocytogenes with an intact leucine-rich repeat region is sufficient to promote internalization. Infect. Immun. 1997, 65, 5309–5319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, L.; Dramsi, S.; Dehoux, P.; Bierne, H.; Lindahl, G.; Cossart, P. InlB: An invasion protein of Listeria monocytogenes with a novel type of surface association. Mol. Microbiol. 1997, 25, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, M.; Copp, J.; Vuga, D.; Marino, M.; Chapman, T.; Van Der Geer, P.; Ghosh, P. GW domains of the Listeria monocytogenes invasion protein InlB are required for potentiation of Met activation. Mol. Microbiol. 2004, 52, 257–271. [Google Scholar] [CrossRef]
- Chalenko, Y.; Kalinin, E.; Marchenkov, V.; Sysolyatina, E.; Surin, A.; Sobyanin, K.; Ermolaeva, S. Phylogenetically Defined Isoforms of Listeria monocytogenes Invasion Factor InlB Differently Activate Intracellular Signaling Pathways and Interact with the Receptor gC1q-R. Int. J. Mol. Sci. 2019, 20, 4138. [Google Scholar] [CrossRef] [Green Version]
- Lecuit, M.; Vandormael-Pournin, S.; Lefort, J.; Huerre, M.; Gounon, P.; Dupuy, C.; Babinet, C.; Cossart, P. A transgenic model for listeriosis: Role of internalin in crossing the intestinal barrier. Science 2001, 292, 1722–1725. [Google Scholar] [CrossRef] [Green Version]
- Pentecost, M.; Kumaran, J.; Ghosh, P.; Amieva, M.R. Listeria monocytogenes internalin B activates junctional endocytosis to accelerate intestinal invasion. PLoS Pathog. 2010, 6, e1000900. [Google Scholar] [CrossRef]
- Chiba, S.; Nagai, T.; Hayashi, T.; Baba, Y.; Nagai, S.; Koyasu, S. Listerial invasion protein internalin B promotes entry into ileal Peyer’s patches in vivo. Microbiol. Immunol. 2011, 55, 123–129. [Google Scholar] [CrossRef]
- Sobyanin, K.; Sysolyatina, E.; Krivozubov, M.; Chalenko, Y.; Karyagina, A.; Ermolaeva, S. Naturally occurring InlB variants that support intragastric Listeria monocytogenes infection in mice. FEMS Microbiol. Lett. 2017, 364, fnx011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabanes, D.; Lecuit, M.; Cossart, P. Animal models of Listeria infection. Curr. Protoc. Microbiol. 2008. [Google Scholar] [CrossRef]
- Camejo, A.; Carvalho, F.; Reis, O.; Leitão, E.; Sousa, S.; Cabanes, D. The arsenal of virulence factors deployed by Listeria monocytogenes to promote its cell infection cycle. Virulence 2011, 2, 379–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecuit, M. Human listeriosis and animal models. Microbes Infect. 2007, 9, 1216–1225. [Google Scholar] [CrossRef] [PubMed]
- Khelef, N.; Lecuit, M.; Bierne, H.; Cossart, P. Species specificity of the Listeria monocytogenes InlB protein. Cell. Microbiol. 2006, 8, 457–470. [Google Scholar] [CrossRef]
- Blanot, S.; Joly, M.M.; Vilde, F.; Jaubert, F.; Clement, O.; Frija, G.; Berche, P. A gerbil model for rhombencephalitis due to Listeria monocytogenes. Microb. Pathog. 1997, 23, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Russ, J.M.; Hutson, A.M.; Montgomery, W.I.; Racey, P.A.; Speakman, J.R. The status of Nathusius’ pipistrelle (Pipistrellus nathusii Keyserling & Blasius, 1839) in the British Isles. J. Zool. 2001, 254, 91–100. [Google Scholar]
- Ciechanowski, M.; Jarzembowski, T. The size and number of harems in the polygynous bat Pipistrellus nathusii (Keyserling and Blasius, 1839) (Chiroptera: Vespertilionidae). Mamm. Biol. 2004, 69, 277–280. [Google Scholar] [CrossRef]
- Dabiri, G.A.; Sanger, J.M.; Portnoy, D.A.; Southwick, F.S. Listeria monocytogenes moves rapidly through the host-cell cytoplasm by inducing directional actin assembly. Proc. Natl. Acad. Sci. USA 1990, 87, 6068–6072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mounier, J.; Ryter, A.; Coquis-Rondon, M.; Sansonetti, P.J. Intracellular and cell-to-cell spread of Listeria monocytogenes involves interaction with F-actin in the enterocytelike cell line Caco-2. Infect. Immun. 1990, 58, 1048–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilney, L.G.; Portnoy, D.A. Actin filaments and the growth, movement, and spread of the intracellular bacterial parasite, Listeria monocytogenes. J. Cell Biol. 1989, 109, 1597–1608. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, J.-L.; Berche, P.; Frehel, C.; Gouln, E.; Cossart, P. Entry of L. monocytogenes into cells is mediated by internalin, a repeat protein reminiscent of surface antigens from gram-positive cocci. Cell 1991, 65, 1127–1141. [Google Scholar] [CrossRef]
- Dramsi, S.; Biswas, I.; Maguin, E.; Braun, L.; Mastroeni, P.; Cossart, P. Entry of Listeria monocytogenes into hepatocytes requires expression of InIB, a surface protein of the internalin multigene family. Mol. Microbiol. 1995, 16, 251–261. [Google Scholar] [CrossRef]
- McNeill, H.; Ozawa, M.; Kemler, R.; Nelson, W.J. Novel function of the cell adhesion molecule uvomorulin as an inducer of cell surface polarity. Cell 1990, 62, 309–316. [Google Scholar] [CrossRef]
- Di Renzo, M.F.; Narsimhan, R.P.; Olivero, M.; Bretti, S.; Giordano, S.; Medico, E.; Gaglia, P.; Zara, P.; Comoglio, P.M. Expression of the Met/HGF receptor in normal and neoplastic human tissues. Oncogene 1991, 6, 1997–2003. [Google Scholar]
- Gloushankova, N.A.; Rubtsova, S.N.; Zhitnyak, I.Y. Cadherin-mediated cell-cell interactions in normal and cancer cells. Tissue Barriers 2017, 5, e1356900. [Google Scholar] [CrossRef] [Green Version]
- Marona, P.; Górka, J.; Kotlinowski, J.; Majka, M.; Jura, J.; Miekus, K. C-Met as a Key Factor Responsible for Sustaining Undifferentiated Phenotype and Therapy Resistance in Renal Carcinomas. Cells 2019, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- Wollert, T.; Pasche, B.; Rochon, M.; Deppenmeier, S.; van den Heuvel, J.; Gruber, A.D.; Heinz, D.W.; Lengeling, A.; Schubert, W.-D. Extending the Host Range of Listeria monocytogenes by Rational Protein Design. Cell 2007, 129, 891–902. [Google Scholar] [CrossRef] [Green Version]
- Niemann, H.H.; Jäger, V.; Butler, P.J.G.; van den Heuvel, J.; Schmidt, S.; Ferraris, D.; Gherardi, E.; Heinz, D.W. Structure of the Human Receptor Tyrosine Kinase Met in Complex with the Listeria Invasion Protein InlB. Cell 2007, 130, 235–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecuit, M.; Dramsi, S.; Gottardi, C.; Fedor-Chaiken, M.; Gumbiner, B.; Cossart, P. A single amino acid in E-cadherin responsible for host specificity towards the human pathogen Listeria monocytogenes. EMBO J. 1999, 18, 3956–3963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, D.L. SARS-CoV-2 and Bats: From Flight to Fighting COVID-19. JACC Basic Transl. Sci. 2020, 5, 545–546. [Google Scholar] [CrossRef] [PubMed]
- Litvin, V.I.; Pushkareva, V.I.; Emel’ianenko, E.N. Biocenosis of the natural foci of sapronotic infections (the results of 15-year observations). Zh. Mikrobiol. Epidemiol. Immunobiol. 2004, 4, 102–108. [Google Scholar]
- Jeffery, S.; Van Der Putten, W.H. Soil Borne Human Diseases. JRC Sci. Tech. Rep. 2011, 49, 37199. [Google Scholar]
- George, D.B.; Webb, C.T.; Farnsworth, M.L.; O’Shea, T.J.; Bowen, R.A.; Smith, D.L.; Stanley, T.R.; Ellison, L.E.; Rupprecht, C.E. Host and viral ecology determine bat rabies seasonality and maintenance. Proc. Natl. Acad. Sci. USA. 2011, 108, 10208–10213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.R.; Won, P.O. Chromosomes of Korean bats. J. Mammal. Soc. Jpn. 1978, 7, 199–203. [Google Scholar]
- Dzuyev, R.I.; Hasнkulova, M.A.; Bogotova, I.H. Features of the chromosome set and measurements of body mediterranean bat (pipisrellus kuhli) in the conditions of forest-steppe zone macrolone the northern central Caucasus. Mod. Probl. Sci. Educ. 2016, 3, 390–398. [Google Scholar]
- Volleth, M.; Bronner, G.; Göpfert, M.C.; Heller, K.G.; Helversen, O.V.; Yong, H.S. Karyotype comparison and phylogenetic relationships of Pipistrellus-like bats (Vespertilionidae; Chiroptera; Mammalia). Chromosom. Res. 2001, 9, 25–46. [Google Scholar] [CrossRef]
- Parida, S.; Domann, E.; Rohde, M.; Müller, S.; Darji, A.; Hain, T.; Wehland, J.; Chakraborty, T. Internalin B is essential for adhesion and mediates the invasion of Listeria monocytogenes into human endothelial cells. Mol. Microbiol. 1998, 28, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Velge, P.; Bottreau, E.; Kaeffer, B.; Pardon, P. Cell immortalization enhances Listeria monocytogenes invasion. Med. Microbiol. Immunol. 1994, 183, 145–158. [Google Scholar] [CrossRef]
- Portnoy, D.A.; Jacks, P.S.; Hinrichs, D.J. Role of hemolysin for the intracellular growth of Listeria monocytogenes. J. Exp. Med. 1988, 167, 1459–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaillard, J.L.; Berche, P.; Mounier, J.; Richard, S.; Sansonetti, P. In vitro model of penetration and intracellular growth of Listeria monocytogenes in the human enterocyte-like cell line Caco-2. Infect. Immun. 1987, 55, 2822–2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortega, F.E.; Koslover, E.F.; Theriot, J.A. Listeria monocytogenes cell-to-cell spread in epithelia is heterogeneous and dominated by rare pioneer bacteria. eLife 2019, 8, e40032. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, J.L. The inlAB locus mediates the entry of Listeria monocytogenes into hepatocytes in vivo. J. Exp. Med. 1996, 183, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Yurkov, S.G.; Kolbasova, O.L.; Zuev, V.V.; Sidorov, S.I.; Kushnir, S.D.; Smyslova, N.Y.U.; Neverovskaya, N.S.; Chermashenceva, N.A.; Prilepskaya, E.P.; Anisimova, L.I. Katalog Kollekcii Kletochnyh Kul’tur VNIIVViM; Rossel’hozakademiya, VNIIVViM: Pokrov, Russia, 2010. [Google Scholar]
- Marchenko, S.; Flanagan, L. Counting Human Neural Stem Cells. J. Vis. Exp. 2007, 7, e262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, C.E.; Hamerton, J.L. A Colchicine, Hypotonic Citrate, Squash Sequence for Mammalian Chromosomes. Stain Technol. 1956, 31, 247–251. [Google Scholar] [CrossRef]
- Rothfels, K.H.; Siminovitch, L. An Air-Drying Technique for Flattening Chromosomes in Mammalian Cells Grown In Vitro. Stain Technol. 1958, 33, 73–77. [Google Scholar] [CrossRef]
- Glaser, P.; Frangeul, L.; Buchrieser, C.; Rusniok, C.; Amend, A.; Baquero, F.; Berche, P.; Bloecker, H.; Brandt, P.; Chakraborty, T.; et al. Comparative genomics of Listeria species. Science 2001, 294, 849–852. [Google Scholar]
- Kibardin, A.; Karpova, T.; Sapenko, T.; Vazquez-Boland, J.A.; Kiselev, S.; Ermolaeva, S. Mammalian peptidoglycan recognition protein TagL inhibits Listeria monocytogenes invasion into epithelial cells. FEMS Immunol. Med. Microbiol. 2006, 46, 284–290. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, D.J.; Klaenhammer, T.R. High- and low-copy-number Lactococcus shuttle cloning vectors with features for clone screening. Gene 1993, 137, 227–231. [Google Scholar] [CrossRef]
- Suárez, M.; González-Zorn, B.; Vega, Y.; Chico-Calero, I.; Vázquez-Boland, J.A. A role for ActA in epithelial cell invasion by Listeria monocytogenes. Cell. Microbiol. 2001, 3, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Ermolaeva, S.; Novella, S.; Vega, Y.; Ripio, M.T.; Scortti, M.; Vázquez-Boland, J.A. Negative control of Listeria monocytogenes virulence genes by a diffusible autorepressor. Mol. Microbiol. 2004, 52, 601–611. [Google Scholar] [CrossRef]
- Chalenko, Y.M.; Sysolyatina, E.V.; Kalinin, E.V.; Sobyanin, K.A.; Ermolaeva, S.A. Natural variants of Listeria monocytogenes internalin B with different ability to stimulate cell proliferation and cytoskeleton rearrangement in HEp-2 cells. Mol. Genet. Microbiol. Virol. 2017, 32, 80–86. [Google Scholar] [CrossRef]
- Sokolovic, Z.; Riedel, J.; Wuenscher, M.; Goebel, W. Surface-associated, PrfA-regulated proteins of Listeria monocytogenes synthesized under stress conditions. Mol. Microbiol. 1993, 8, 219–227. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immunization | Week | Dose (mg) | Adjuvant | Mode of Administration |
|---|---|---|---|---|
| 1st | 0 | 0.5 | - | Iv |
| 0.375 | CFA | Sc | ||
| 0.375 | CFA | Im | ||
| 2nd | 4 | 0.25 | - | Iv |
| 0.75 | IFA | Sc | ||
| 3rd | 8 | 0.25 | - | Iv |
| 0.75 | IFA | Sc | ||
| 4th | 10 | 0.25 | - | Iv |
| 0.75 | IFA | Sc | ||
| 12 | Sampling | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Povolyaeva, O.; Chalenko, Y.; Kalinin, E.; Kolbasova, O.; Pivova, E.; Kolbasov, D.; Yurkov, S.; Ermolaeva, S. Listeria monocytogenes Infection of Bat Pipistrellus nathusii Epithelial cells Depends on the Invasion Factors InlA and InlB. Pathogens 2020, 9, 867. https://doi.org/10.3390/pathogens9110867
Povolyaeva O, Chalenko Y, Kalinin E, Kolbasova O, Pivova E, Kolbasov D, Yurkov S, Ermolaeva S. Listeria monocytogenes Infection of Bat Pipistrellus nathusii Epithelial cells Depends on the Invasion Factors InlA and InlB. Pathogens. 2020; 9(11):867. https://doi.org/10.3390/pathogens9110867
Chicago/Turabian StylePovolyaeva, Olga, Yaroslava Chalenko, Egor Kalinin, Olga Kolbasova, Elena Pivova, Denis Kolbasov, Sergey Yurkov, and Svetlana Ermolaeva. 2020. "Listeria monocytogenes Infection of Bat Pipistrellus nathusii Epithelial cells Depends on the Invasion Factors InlA and InlB" Pathogens 9, no. 11: 867. https://doi.org/10.3390/pathogens9110867